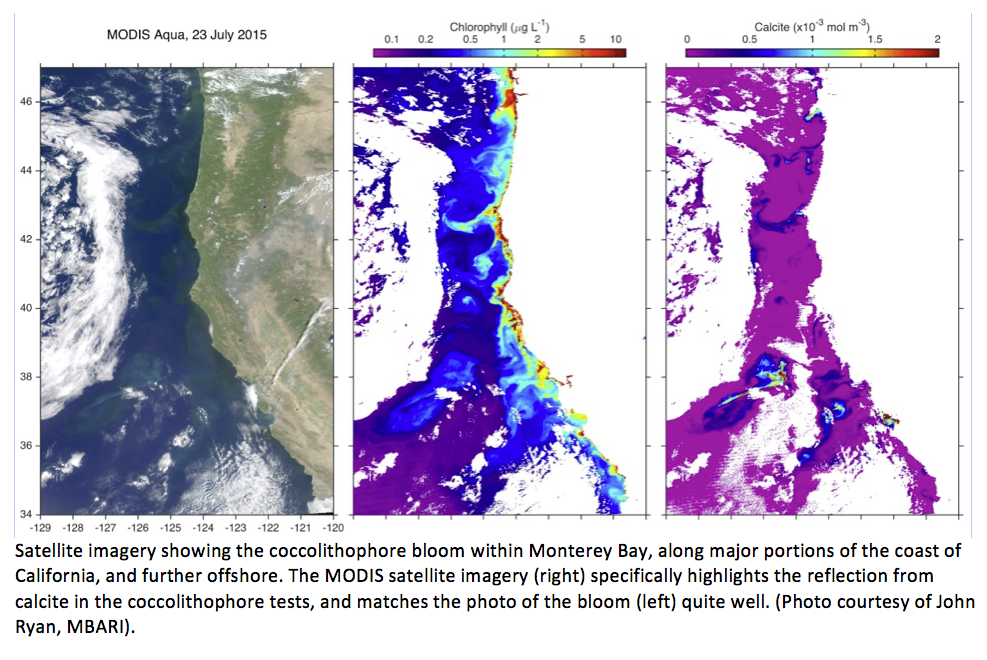
 During the middle weeks of July 2015 the normally deep blue waters of Monterey Bay turned a vibrant turquoise color. This was caused by a phytoplankton bloom, a tremendous increase in the abundance of a single-celled alga known as a coccolithophore. Coccolithophores reflect light back as greenish-blue hues, and when high densities occur these light-scattering properties alter the ocean’s appearance. Although the ocean appears turquoise from above, the water within a coccolithophore bloom actually looks milky-white from underneath, as divers will confirm.
During the middle weeks of July 2015 the normally deep blue waters of Monterey Bay turned a vibrant turquoise color. This was caused by a phytoplankton bloom, a tremendous increase in the abundance of a single-celled alga known as a coccolithophore. Coccolithophores reflect light back as greenish-blue hues, and when high densities occur these light-scattering properties alter the ocean’s appearance. Although the ocean appears turquoise from above, the water within a coccolithophore bloom actually looks milky-white from underneath, as divers will confirm.
These coccolithophore blooms occur throughout the world, and earlier this summer along California’s coast near Santa Barbara a coccolithophore bloom turned waters turquoise. Phytoplankton researchers from UC Santa Barbara took water samples within Monterey Bay in July 2015 and confirmed that the turquoise color of the water was indeed caused by a large bloom of coccolithophores, estimating densities of up to 1 million/cubic liter within Monterey Bay. The bloom persisted for about three weeks but diminished toward the end of July as minor upwelling and advection diluted the bloom. Besides the earlier bloom near Santa Barbara and more recently within Monterey Bay, scientists have identified additional coccolithophore blooms further north and offshore from the coast.
 As a single-celled alga, coccolithophores are part of the world’s most abundant type of primary producers, which form the basis of pelagic marine food webs. Although other phytoplankton contain similar photosynthetic pigments, coccolithophores are unique in having calcite disks (coccoliths) composed of calcium carbonate. These calcium carbonate plates are arranged into a coccosphere that are shed off and contribute to the reflective properties of the bloom. The evolutionary origins of the coccosphere are uncertain but its true adaptive qualities may have served as protection from predation or additional environmental pressures such as osmotic and chemical shock or UV exposure. Other phytoplankton, including certain diatoms and dinoflagellates, are also encased in a shell but they lack the optical properties of the coccolith.
As a single-celled alga, coccolithophores are part of the world’s most abundant type of primary producers, which form the basis of pelagic marine food webs. Although other phytoplankton contain similar photosynthetic pigments, coccolithophores are unique in having calcite disks (coccoliths) composed of calcium carbonate. These calcium carbonate plates are arranged into a coccosphere that are shed off and contribute to the reflective properties of the bloom. The evolutionary origins of the coccosphere are uncertain but its true adaptive qualities may have served as protection from predation or additional environmental pressures such as osmotic and chemical shock or UV exposure. Other phytoplankton, including certain diatoms and dinoflagellates, are also encased in a shell but they lack the optical properties of the coccolith.
In contrast to the nutrient-rich waters that diatoms thrive in, the blooms of coccolithophores are relatively chlorophyll-a poor and tend to appear after diatom blooms, taking advantage of a shallow mixed, upper-layer after reformation of the summer thermocline. This upper layer traps light and heat and allows light to refract intensely through the bloom, generating reflective wavelengths of a turquoise color. Coccolithophores fill a unique ecological niche that allows for major blooms to seemingly appear from nowhere based on small “seed” populations acting as propagules. Coccolithophores can rapidly reproduce by the process of binary fission reaching levels as high as 1,000,000 per liter.
 Most evidence for coccolithophore blooms come from satellite imagery, and are verified by analyzing in situ water samples. Although there are over ninety recognized species of coccolithophores in the Pacific Ocean the species that blooms most frequently and intensely is Emiliania huxleyi so it is likely that the bloom in the bay is of this species. Emiliania huxleyi is a cosmopolitan species found throughout the oceans from the tropics to the subartic regions. Like other wide-spread species, the broad habitat range of E. huxelyi indicates it became a cosmopolitan species because it can tolerate a wide variety of environmental conditions such as temperature, salinity, or other water quality conditions. However, specific common water quality parameters have been identified as triggering a bloom. Most commonly this includes nutrient-poor waters, high light, and strong ocean stratification as mentioned. Additionally, other ambient conditions include low silicate levels, an unusually high ratio of nitrogen to phosphate, low dissolved carbon dioxide, and a high carbonate saturation rate. The types of zooplankton grazers in the area may also limit or allow the coccolithophores to bloom.
Most evidence for coccolithophore blooms come from satellite imagery, and are verified by analyzing in situ water samples. Although there are over ninety recognized species of coccolithophores in the Pacific Ocean the species that blooms most frequently and intensely is Emiliania huxleyi so it is likely that the bloom in the bay is of this species. Emiliania huxleyi is a cosmopolitan species found throughout the oceans from the tropics to the subartic regions. Like other wide-spread species, the broad habitat range of E. huxelyi indicates it became a cosmopolitan species because it can tolerate a wide variety of environmental conditions such as temperature, salinity, or other water quality conditions. However, specific common water quality parameters have been identified as triggering a bloom. Most commonly this includes nutrient-poor waters, high light, and strong ocean stratification as mentioned. Additionally, other ambient conditions include low silicate levels, an unusually high ratio of nitrogen to phosphate, low dissolved carbon dioxide, and a high carbonate saturation rate. The types of zooplankton grazers in the area may also limit or allow the coccolithophores to bloom.
Coccolithophore populations do not appear along California’s coast every summer, but the 2015 summer has seen unusually warm coastal waters. The recent harmful algae bloom (HAB) of a different phytoplankton species that appeared in Monterey Bay was part of a massive HAB stretching along major portions of the west coast and is thought to be linked to an expanse of warm water that has settled offshore.
 Relatively warm water this year and the extreme stratification of bay waters are an ideal habitat for coccolithophore blooms to occur. With increasing ocean temperatures predicted by climate change it will be important to continuously monitor the frequency and extent of coccolithophore blooms and the possible effects the blooms have on the environments throughout Monterey Bay and the California coast.
Relatively warm water this year and the extreme stratification of bay waters are an ideal habitat for coccolithophore blooms to occur. With increasing ocean temperatures predicted by climate change it will be important to continuously monitor the frequency and extent of coccolithophore blooms and the possible effects the blooms have on the environments throughout Monterey Bay and the California coast.
10 interesting facts about coccolithophores:
- The first reported blooms of Emiliania huxleyi came from fjords on the western coast of Norway in the 1950’s.
- A coccolithophore bloom is defined by concentrations of at least 1,000,000 algal cells/L water.
- Coccolithophore blooms act as a significant carbon sink, and are thought to be important in the global carbon cycle.
- Coccolithophore blooms will commonly cover extraordinarily large areas, for example, over 200,000 sq. km in the eastern Bering Sea.
- The most intense bloom ever recorded was during the summer of 1955 in fjord in Norway and had coccolithophore concentrations of 150,000,000 huxleyi cells per liter.
- Emiliania huxleyi is an unusual coccolithophore in that is overproduces coccoliths during blooms, which are shed off and increase the light-scattering properties of a bloom that make the water appear turquoise.
- Emiliania huxleyi and other coccolithophores produce large quantities of dimethyl sulfide (DMS), the most abundant biological sulfur compound emitted to the atmosphere.
- Limiting silicate in the water is an important environmental condition for Emiliania huxleyi blooms because it limits the ability of its main phytoplankton competitor, the diatom, from blooming.
- Emiliania huxleyi produces very long-chain n-alkenes and n-alkenones that may be a useful biological marker for species identification.
- The White Cliffs of Dover along the coast of southern England are composed of the chalky coccolithophore shells compacted into the sediment long ago.
Some good websites about coccolithophores:
http://www.noc.soton.ac.uk/soes/staff/tt/eh/index.html
http://ina.tmsoc.org/Nannotax3/index.php?dir=Coccolithophores
List of References:
Egge JK, Aksnes, DL. 1992. Silicate as regulating nutrient in phytoplankton competition. Marine Ecology Progress Series, 83 (2): 281–289, doi:10.3354/meps083281.
Okada HS et al. 1973. The distribution of oceanic coccolithophores in the Pacific. Deep Sea Research and Oceanographic Abstracts, 20 (4): 355–374, doi:10.1016/0011-7471(73)90059-4.
Smith HEK, et al. 2012. Predominance of heavily calcified coccolithophores at low CaCO3 saturation during winter in the Bay of Biscay. Proceedings of the National Academy of Sciences, 109 (23): 8845–8849, doi:10.1073/pnas.1117508109.
Tyrrell T, and Merico A. 2004. Emiliania huxleyi: bloom observations and the conditions that induce them. In Coccolithophores (pp. 75-97). Springer Berlin Heidelberg.
Volkman JK, Eglinton G, Corner ED, and Forsberg TEV. 1980. Long-chain alkenes and alkenones in the marine coccolithophorid Emiliania huxleyi. Phytochemistry, 19(12), 2619-2622.
Wolfe GV, and Steinke M. 1996. Grazing-activated production of dimethyl sulfide (DMS) by two clones of Emiliania huxleyi. Limnology and Oceanography, 41(6), 1151-1160.